
- Модельные системы в нейроэтологии животное
- Поведение
- Нейронные Субстраты
- Картинка предоставлена Комиссия по рыболовству в Великих озерах , Фигура миномоза от Grillner...
- Ресурсы миноги WWW:
Модельные системы в нейроэтологии животное
Наша команда-партнер Artmisto

Морская минога ( Petromyzon marinus), прикрепленная к хозяину
Североамериканцы наиболее знакомы с Морской миногой как с незваным паразитом, который почти полностью уничтожил всех местных рыб Верхних Великих озер после их проникновения через Уэллендский канал в 1921 году. С тех пор многие поклонники преодолели эту дурную славу (и их отсутствие каких-либо искупительных эстетических качеств) благодаря стараниям шведского Стена Грилнера и других нейробиологов за последние два десятилетия, которые сделали его очень важной модельной системой для понимания передвижения позвоночных.
Морская минога может жить в океане или в пресной воде, и идти вверх по рекам, чтобы нереститься. Взрослая морская минога может достигать 120 см (47 дюймов); Великие озера, не имеющие выхода к морю, P. marinus вырастают до 64 см (25 дюймов). Они весят 900-1400 грамм (2-3 фунта) и имеют два спинных плавника и без чешуи. У них рот с острыми зубцами с сосущим диском. Рот вспыхивает, когда они прикрепляются к своим хозяевам с помощью всасывания, а затем их зубец ранит в организм хозяина рану, которая остается открытой антикоагулянтом, удобно генерируемым буккальной железой. Они выживают в жидкостях организма своего хозяина, часто до тех пор, пока хозяин не умрет. Форма личинок живет в грязи в течение 5 лет, а затем превращается во взрослую особь (контроль над популяциями миног в Великих озерах осуществляется с помощью ламприцида, который убивает личинок).
Миноги происходят от одной из старейших существующих групп позвоночных, отошедших от других позвоночных около 500 миллионов лет назад до того, как появилась отрасль, которая привела к появлению современных рыб, и оставались несколько «первичными» с точки зрения ряда мер сложности позвоночных. В нейробиологическом отношении они представляют собой прото-позвоночных с нервной системой, состоящей из стандартных типов и структур клеток позвоночных, но на порядок меньше нейронов, чем у современных позвоночных. Нервная система миноги также достаточно устойчива, что позволяет проводить длительные частичные эксперименты на животных, которые обычно используют нейробиологи-беспозвоночные, и, как правило, очень трудны для современных позвоночных.
Древо жизни: Hyperoartia
Поведение
Простота и надежность нервной системы морских миног, наряду с бесчеловечной формой передвижения (движение, перемещая свое тело вперед и назад по воде), не усложняемой грудными плавниками и другими достижениями современной локомоции рыбы, сделали ее идеальной модельной системой. для изучения локомоции позвоночных. Миноги плавают, распространяя направленную вбок волну по их угрюмому телу. Амплитуда этой волны увеличивается к хвосту. Чем быстрее волна распространяется по телу, тем быстрее минога плавает. Рулевое управление достигается путем сгибания тела в нужном направлении. Один полный цикл плавания вперед и назад показан на рисунке ниже. Общая продолжительность этого цикла варьируется от 4 секунд до 100 миллисекунд (от 0,25 до 10 Гц) с длиной волны, немного меньшей длины тела. Фильм QuickTime об их методе передвижения можно найти на WWW-страница Джозефа Айера о волнистом роботе на основе миног ,
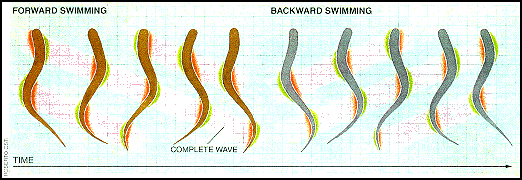
Нейронные Субстраты
За последние 15 лет был достигнут быстрый прогресс в понимании нейронных основ локомоции миног. Исследования проводились на многих уровнях - субклеточном, клеточном, сегментарном, мультисегментном, вычислительном и поведенческом.
Спинной мозг состоит из около 100 сегментов. Каждый сегмент содержит около 1000 нейронов. Они получают иннервацию от глутаматергических ретикулоспинальных нейронов в заднем и среднем ромбэнцефальных ядрах. Эта нисходящая проекция - это то, что инициирует локомоцию, и может также обеспечить тоническое движение. Они, в свою очередь, возбуждаются ростральными структурами головного мозга и несколькими различными типами сенсорных раздражителей, такими как вестибулярная и боковая линия (люди-рыбки будут удивлены, узнав, что на боковой линии миноги есть не только невромасты, которые чувствуют движение воды, но и фоторецепторы на коже области хвоста, которая иннервируется нервами боковой линии туловища. Deliagina и др. показали, что вспышка света на этих датчиках может инициировать локомоцию. Также очень интересным является обнаружение Норткаттом, что минога имеет ампуллярные электрорецепторы). Направленная вбок волна, которая распространяется вдоль тела, обусловлена чередующейся активацией мотонейронов на левой и правой стороне каждого сегмента в сочетании с последовательной активацией заднего (для прямого плавания) или переднего (для обратного плавания) сегментов с фазой задержка.
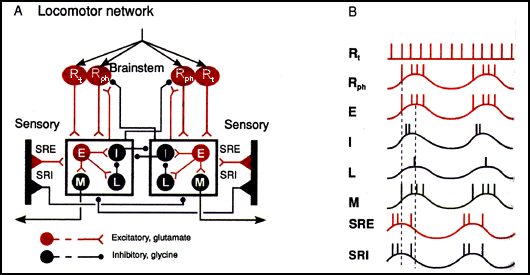
Схема расположения одного из ок. 100 сегментов опосредуют плавание миноги вместе с иннервацией ствола мозга. Синапсы, оканчивающиеся или начинающиеся на кадрах блока, указывают на эффект, общий для всех нейронов в блоке. Заполненные кружки представляют популяции нейронов, а не отдельные единицы. Вертикальные черные полосы - это рецепторы растяжения, расположенные на боковом крае спинного мозга.
тонически (непрерывно) активные ретикулярные нейроны ствола мозга R ph: физически (прерывисто) активные ретикулярные нейроны ствола мозга E: возбуждающие сегментарные межнейроны I: ингибирующие сегментарные межнейроны L: латеральные тормозные интернейроны M: мотонейроны SRE: возбудительные нервные растяжения : ингибирующие нейроны рецептора растяжения
Краткое описание схемы:
(A) Ретикулоспинальные глутаматергические нейроны ствола мозга R t и R ph становятся возбужденными и активируют первую сегментарную нейронную сеть в спинном мозге миноги, возбуждая группы нейронов E , I , L и M на одной стороне (далее именуемые как « активная сторона ") сети. E нейроны возбуждают все нейроны в коробке: глицинергические ингибирующие интернейроны I, которые пересекают срединную линию и ингибируют все нейроны на другой стороне сети, латеральные интернейроны L, которые ингибируют I интернейрон, и холинергические M мотонейроны, которые сжимают плавательные мышцы. С задержкой в 1% (поскольку это 1 из 100 сегментов, составляющих длину шнура) от общего времени цикла, следующий сегмент в направлении распространения волны возбуждается.
По мере изгиба рыбы нейроны рецепторов растяжения SRE и SRI на стороне активного сегмента деактивируются, тогда как нейроны на противоположной стороне возбуждаются растяжением, вызванным сокращением мышц на активной стороне. Таким образом, SRI неактивной стороны начинают ингибировать все нейроны в боксе на противоположной активной стороне, а SRE неактивной стороны начинают возбуждать все нейроны в боксе на неактивной стороне.
Инициирование активности на стороне, противоположной активной в настоящее время стороне, является, главным образом, результатом дезингибирования нейронов на неактивной стороне от нейронов SRI активной стороны в сочетании с фоновым возбуждающим импульсом.
(B) Диаграмма, схематично показывающая отношения между основными группами нейронов на одной стороне сегментарной нейронной сети.
Более точное разрешение
Чтобы понять некоторую сложную динамику сети одного сегмента, представьте, что возбуждаете одну сторону сети агонистом рецептора NMDA (который запускает циклы плавания 0,1-3 Гц) или агонистом рецептора AMPA / каината (инициирует циклы плавания 1-8 Гц). Это приведет к тому, что эта половина будет активной, а мотонейроны сжимают миотомы, которые они иннервируют на этой стороне; активация другой стороны будет ингибироваться глицинергическими ингибиторными интернейронами I активной стороны, которые пересекают срединную линию. Это несмотря на фоновое возбуждение от R t нейронов. Если ничего не изменится, минога будет иметь постоянную асимметричную активность. То, как сеть выпрыгивает из этого состояния, похожего на захват, является полезным способом приблизиться к системе.
Одним из способов является сенсорный, уже описанный выше: расширение неактивной стороны заставляет нейроны SRI ингибировать активную сторону, а нейроны SRE активируют неактивную сторону, которая в сочетании с фоновым возбуждением становится активной.
Другой способ (который в действительности действует совместно с механизмом прекращения сенсорного взрыва) - это электрохимическая динамика нейронов с каждой стороны. Общим концом этих механизмов является манипулирование активированным кальцием током калиевого канала (IKca). Когда этот исходящий ток включен, ячейка переполяризована и прекратит срабатывание потенциалов действия.
Один из механизмов, который я опишу, это прерывание через окончание того, что называется потенциалом плато NMDA. Как упоминалось ранее, активация сегментарной сети с помощью агониста NMDA вызовет частоты всплесков, которые соответствуют медленному плаванию. В клетках с NMDA-рецепторами достаточное возбуждение приводит к активации зависимых от напряжения каналов NMDA, что приводит к длительной деполяризации (потенциал плато). В E-ячейке это привело бы к постоянному возбуждению активной стороны. Но поскольку канал NMDA позволяет кальцию поступать, кальций накапливается в клетке. В конечном итоге это включает IKca и потенциал плато заканчивается. IKca медленно инактивируется, что приводит к длительной гиперполяризации (sAHP) в клетке. Это не только прекращает активность на одной стороне, но также приводит к адаптации частоты скачка, так как sAHP могут накапливаться с последовательными потенциалами действия, что приводит к более длительным периодам отсутствия огня, несмотря на продолжающийся ввод. Постингибирующий отскок неактивной стороны часто достигается комбинацией фоновой активации AMPA / kainate или NMDA в сочетании с включением тока низкого напряжения с активированным кальцием (LVA), который включается только так же, как нейрон. приближается к порогу потенциала действия из ранее заблокированного состояния. Хотя это обычно действует, чтобы дать неактивной стороне дополнительный импульс для начала стрельбы после дезингибирования с противоположной стороны, это также может способствовать IKca и последующей реполяризации.
Модуляция этой динамики достигается с помощью 5-HT (серотонина), GABAb, mGluR и пептидергических систем. Например, 5-НТ и допамин хранятся в средней линии системы нейронов позвоночника. Это действует на дендриты сегментарной сети. 5-HT снижает проводимость IKca, влияя на потенциал плато sAHP и NMDA. Это вызовет более длительные всплески и повлияет на межсегментную задержку фазы. Дофамин уменьшает токи кальция, а также способствует в IKca. Дополнительные механизмы пресинаптической модуляции управляют различными нейронами в сегментарных сетях, поэтому они нечувствительны к супраспинальному вводу во время определенных фаз цикла активации / инактивации, что дает меру локального контроля.
В настоящее время динамика отдельных предметов лучше известна эмпирически, чем многосегментная координация. Известно, что при регулярном плавании вперед происходит постоянная фазовая задержка в 1% между смежными сегментами в рострально-каудальном направлении. Изолированный спинной мозг демонстрирует ту же картину при активации агонистами глутаматных рецепторов. Однако это отставание является переменным и может варьироваться от + 2% для плавания вперед и до -1% для плавания назад. Моделирование в компьютерных исследованиях показало, что эти фазовые задержки могут быть учтены с учетом понятной в настоящее время схемы сегментарной и межсегментной связности с использованием простых нейронных механизмов, но нет единого мнения относительно того, какая модель, если таковая имеется, соответствует реальности in vivo .
В общем, как и в случае с динамикой стоматогастрального ганглия, слова быстро дают сбои в понимании происходящего. Как Grillner и соавт. в своем обзоре TINS '95, мотивируя подход к моделированию, говорится: «К сожалению, наш мозг не может справиться с этим типом динамического процесса, в котором одновременно играют роль несколько факторов». Орджан Экеберг и др. Разработали некую очень захватывающую вычислительную нейронауку, которая создала интегрированную трехмерную нейромеханическую модель локомоции миног, которая сочетает в себе биофизически реалистичные нейроны с имитированными миотомами и водой и приводит к реалистичным образам виртуального движения миноги. (см. Ekeberg et al. '95). Один интересный вопрос для интегративных нейробиологов заключается в том, могут ли такие открытия оставить человеческое механистическое (хотя и не целостное неформальное) понимание позади, так же, как это делают сгенерированные компьютером доказательства теорем (см. Ссылки на специальные темы в конце этой страницы, чтобы следовать этой теме) ,
Картинка предоставлена Комиссия по рыболовству в Великих озерах , Фигура миномоза от Grillner 95 года. Принципиальная схема Grillner et al. '95. Резюме основано на Grillner et al. 95, Grillner et al. '91, и Ekeberg et al. '95.
Избранные ссылки
Ранняя классика:
Ровайнен, CM (1974a) Синаптические взаимодействия выявленных нервных клеток в спинном мозге морской миноги. J. Сравнительная неврология : 154: 189-206.
Примечание: нажмите на ссылку автора, чтобы увидеть анализ цитирования этой статьи .
Ровайнен С.М. (1974b) Синаптические взаимодействия ретикулоспинальных нейронов и нервных клеток в спинном мозге морской миноги. J. Сравнительная неврология : 154: 207-223.
Grillner, S., McClellan, A. и Perret, C. (1981). Улавливание генераторов спинного мозга для плавания механочувствительными элементами в миноге спинного мозга in vitro. Исследование мозга 217: 380-386.
Grillner S. (1985). Нейробиологические основы ритмических двигательных актов у позвоночных. Science 228: 143-49.
Отзывы:
Nieuwenhuys, R. (1977) Мозг миноги в сравнительной перспективе. Летопись Нью-Йоркской академии наук 299: 97-145.
Grillner, S. (1996) Нейронные сети для передвижения позвоночных. Scientific American , 274: 64-69. Полный текст статьи с рисунками доступен на Cure Paralysis Now WWW сайт ,
Grillner S., Deliagina, T, Ekeberg, ..., Manira, A. El, Hill, RH, Lansner, A., Orlovsky, GN, P. Wallén (1995) Нейронные сети, которые координируют локомоцию и ориентацию тела в минога. Тенденции в Neuroscience 18 (6): 270-279.
Grillner S., Wallén P., Brodin L. and A. Lansner (1991). Нейронная сеть, генерирующая локомоторное поведение миног: схема, передатчики, свойства мембраны и моделирование. Ежегодный обзор нейронауки 14: 169-199.
Некоторые последние достижения:
Бьюкенен, JT и DR. Макферсон (1995). Нейронная сеть для локомоции спинного мозга миноги - свидетельство вовлечения спаечных интернейронов. Журнал физиологии (Париж) 89 (4-6): 221-233.
Ekeberg, ..., Lansnser, A. and S. Grillner (1995). Нейронный контроль плавания рыб изучен с помощью численного моделирования. Адаптивное поведение 3: 363-384. Обсуждается интегрированная нейромеханическая модель, упомянутая выше.
(Для другой интересной интегрированной вычислительной модели плавания и поведения рыб см .: Terzopoulos D., Tu X, Grzeszczuk R. (1995). Искусственные рыбы: автономное передвижение, восприятие, поведение и обучение в моделируемом физическом мире. Искусственная жизнь : 1: 327-351.)
Jung R., Kiemel T. and AH Cohen (1996). Динамическое поведение нейросетевой модели локомоторного управления у миног. Журнал нейрофизиологии 75 (3): 1074-1086.
Schotland JL, Shupliakov O., Grillner S и L. Brodin (1996). Синаптические и несинаптические моноаминергические нейронные системы в спинном мозге миног. Журнал сравнительной неврологии 372 (2): 229-244.
Vinay, L., Barthe, JY и S. Grillner (1996). Центральная модуляция рецепторов растяжения при фиктивной локомоции у миног. Журнал нейрофизиологии 72 (2): 1224-1235.
Специальная тема: сложность против редукционистского понимания
Андерсон, PW (1972) Больше другое. Science 177: 393-396.
Баллок, TH (1983). Эпилог: нейробиологические корни и нейроэтологические ростки. В кн . : Нейроэтология и поведенческая физиология , Хьюбер Ф. и Х. Маркль, ред. Springer-Verlag 1983.
De Millo R., Lipton RJ и AJ Perlis (1979). Социальные процессы и доказательства теорем и программ. Сообщения ACM 22: 271-280.
Гарфинкель Д. (1980). Компьютерное моделирование, сложные биологические системы и их упрощения. Американский журнал физиологии 239 (регулятивная интегративная сравнительная физиология 8): R1-R6.
Selverston, AI (1980). Понятны ли генераторы центрального шаблона? Поведенческие и мозговые науки 3, 535-571.
Вайнберг Г.М. (1975). Введение в общее мышление систем . Нью-Йорк: Уайли.
Ресурсы миноги WWW:
Полный текст научной статьи С. Грилнера «Нейронные сети для управления движением»
Эксперименты с нейромеханической моделью миноги
Агната ссылки
Волнообразный робот на основе миног
Естественная история миноги
Программа контроля миног Великих озер
Fish Locomotion Библиография и рефераты



